La histología vegetal trata del estudio de todos los tejidos orgánicos propios de las plantas.
En una planta vascular existen tejidos diferenciados de acuerdo a la función que desempeñan: tejidos de crecimiento (meristemos), protectores (epidermis y peridermis), fundamentales (parénquima), de sostén (colénquima y esclerénquima), conductores (floema y xilema).
En una planta vascular existen tejidos diferenciados de acuerdo a la función que desempeñan: tejidos de crecimiento (meristemos), protectores (epidermis y peridermis), fundamentales (parénquima), de sostén (colénquima y esclerénquima), conductores (floema y xilema).
Meristemos: características citológicas
Los meristemos son los responsables del crecimiento permanente de las plantas y están presentes durante toda la vida de éstas. Las células meristemáticas presentan las características citológicas de las células indiferenciadas. Son pequeñas, isodiamétricas y tienen una pared celular primaria delgada. Su citoplasma contiene características propias, como abundantes ribosomas, un retículo endoplasmático rugoso escaso, el complejo de Golgi muy desarrollado para fabricar los componentes de la pared celular, numerosos proplastidios, muchas y pequeñas vacuolas y un protoplasma desprovisto de inclusiones.
Se localizan en las semillas, en los ápices de las raíces y los tallos, en las yemas y también en el interior del tallo o tronco. Frecuentemente, cuando se observa al microscopio, se puede ver que algunas (o muchas) de sus células se encuentran en división.
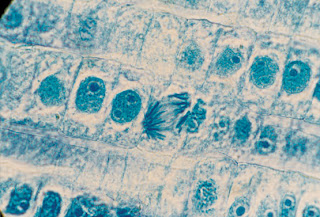
Se localizan en las semillas, en los ápices de las raíces y los tallos, en las yemas y también en el interior del tallo o tronco. Frecuentemente, cuando se observa al microscopio, se puede ver que algunas (o muchas) de sus células se encuentran en división.

CÉLULAS MERISTEMÁTICAS

Clasificación de los meristemos
Los meristemas se clasifican por su localización en el cuerpo de la planta en:
Apicales
Laterales
Intercalares
Teniendo en cuenta además el tiempo de aparición del meristema, se los clasifica en primarios y secundarios. Los meristemas apicales son primarios, mientras los laterales (cámbium y felógeno), son secundarios.
Meristemos apicales:
Los primeros mersitemos en aparecer durante el desarrollo del cuerpo vegetativo de una planta vascular están localizados en la punta de tallos y raíces. debido a su localización, estos meristemos son llamados meristemos apicales. Todos los tejidos meristemáticos primarios y por lo tanto todos los tejidos primarios de la planta se originan a partir del meristemo apical de la raíz o del meristemo apical del brote. El meristemo apical de la raíz normalmente esta cubierto por una estructura de células diferenciadas que lo protege, conocida como cofia. El meristemo apical del tallo (o yema terminal) puede estar desnudo o cubierto por hojas.


Los primeros mersitemos en aparecer durante el desarrollo del cuerpo vegetativo de una planta vascular están localizados en la punta de tallos y raíces. debido a su localización, estos meristemos son llamados meristemos apicales. Todos los tejidos meristemáticos primarios y por lo tanto todos los tejidos primarios de la planta se originan a partir del meristemo apical de la raíz o del meristemo apical del brote. El meristemo apical de la raíz normalmente esta cubierto por una estructura de células diferenciadas que lo protege, conocida como cofia. El meristemo apical del tallo (o yema terminal) puede estar desnudo o cubierto por hojas.
Meristemos laterales:
Producen el engrosamiento de los tallos y las raíces. Responsables del crecimiento radial (secundario). Están distribuidos por toda la planta.Dan lugar a xilema, floema y parénquima secundario (cámbium) y a parénquima cortical y suber (felógeno), y contribuyen al engrosamiento de tallos y raíces por formación de capas concéntricas nuevas que dan lugar además a un engrosamiento de los ejes. Sus células recuperan su capacidad meristemática y comienzan a dividirse formando nuevas células, dando lugar a un crecimiento en grosor en tallos y raíces de plantas leñosas.

Producen el engrosamiento de los tallos y las raíces. Responsables del crecimiento radial (secundario). Están distribuidos por toda la planta.Dan lugar a xilema, floema y parénquima secundario (cámbium) y a parénquima cortical y suber (felógeno), y contribuyen al engrosamiento de tallos y raíces por formación de capas concéntricas nuevas que dan lugar además a un engrosamiento de los ejes. Sus células recuperan su capacidad meristemática y comienzan a dividirse formando nuevas células, dando lugar a un crecimiento en grosor en tallos y raíces de plantas leñosas.
Meristemos intercalares
Se encuentran intercalados entre tejidos no meristemáticos. Los mejor conocidos son aquellos localizados en los entrenudos (principalmente la base del entrenudo) y en la vaina de las monocotiledóneas.
Los meristemas intercalares son zonas de tejido primario en crecimiento activo, situadas entre regiones de tejidos más o menos diferenciadas.
Meristemas intercalares en gramínea | |
Un ejemplo muy conocido son los meristemas que se hallan en los entrenudos y en las vainas foliares de muchas monocotiledóneas, son los responsables del crecimiento en altura de la planta. |  |
Concepto y clasificación de los tejidos
Tejido es una asociación de células que tienen un origen común y que en conjunto cumplen las mismas funciones. Se forman a partir de células apicales o conos vegetativos.
Tejidos de sostén
El colénquima es un tejido vivo formado por un solo tipo celular, la célula colenquimática. Presenta una gruesa pared celular primaria caracterizada por engrosamientos distribuidos de manera desigual y esto confiere al tejido gran fuerza de tensión y resistencia al estrés mecánico. Las células colenquimáticas, al igual que las células parenquimáticas, son capaces de reanudar una actividad meristemática gracias a que sus paredes celulares son primarias y no lignificadas, a pesar de su grosor. Es un tejido poco extendido en el cuerpo de las plantas ya que, por lo general, no está presente en las raíces ni tampoco en estructuras con crecimiento secundario, donde es sustituido por el esclerénquima.
Colénquima angular de una hiedra.

Colénquima laminar en el tallo de un saúco.

El esclerénquima, a diferencia del colénquima, presenta dos tipos de células con pared celular engrosada, pero ésta es secundaria y lignificada en las células maduras. Las células esclerenquimáticas maduras no contienen protoplasma y son células muertas. Gracias a la estructura de sus paredes celulares el esclerénquima tiene una función muy importante en el soporte de los órganos que han dejado de alargarse. Protegen las partes más blandas de las plantas y más vulnerables a estiramientos, pesos, presiones y flexiones. Por eso aunque está distribuido por todo el cuerpo de las plantas, ya sean estructuras con crecimiento primario o secundario, es más abundante en tallos y hojas que en raíces. Este tejido es complejo y los dos tipos de células que lo componen se distinguen principalmente por su forma, su origen y su localización. Un tipo son las fibras, células alargadas y fusiformes, y el otro las esclereidas, que son células variadas en su forma pero típicamente más isodiamétricas que las fibras.
Fibras de esclerénquima del maíz.

Fibras de esclerénquima del maíz.

Tejidos conductores
La característica más llamativa que distingue a las plantas vasculares de las no vasculares es la presencia en las primeras de tejidos vasculares especializados en la conducción de agua y sustancias inorgánicas y orgánicas. Estos tejidos son el xilema y el floema.
Los tejidos conductores son complejos y están formados por distintos tipos celulares, la mayor parte de los cuales se originan de las mismas células meristemáticas
En el XILEMA, también llamado leño, nos encontramos cuatro tipos celulares principales: las traqueidas y los elementos de los vasos, que son las células conductoras o traqueales, las células parenquimáticas y las fibras de esclerénquima, que funcionan como células de almacenamiento y sostén, respectivamente.


Las traqueidas son células alargadas, estrechas y fusiformes. El agua circula por ellas y pasa de unas a otras vía simplasto atravesando las punteaduras areoladas, que se encuentran en sus paredes laterales.
Los elementos de los vasos son células de mayor diámetro y más achatadas que las traqueidas. Se unen longitudinalmente unas a otras para formar tubos llamados vasos o tráqueas.
El FLOEMA, llamado líber o tejido criboso, está formado por más tipos celulares que el xilema. Los elementos conductores son la célula cribosa y los tubos cribosos y dentro de los elementos no conductores se encuentran las fibras de esclerénquima y las células parenquimáticas. Las células parenquimáticas pueden ser típicas y especializadas, acompañando estas últimas a los elementos conductores.
Las células cribosas son largas y de extremos puntiagudos, comunicándose entre sí lateralmente mediante grupos de campos de poros primarios que forman las áreas cribosas.

Tejidos de proteccion
Los tejidos de protección forman el límite externo de las plantas y se encuentran en contacto con el medio ambiente. Hay dos tipos dependiendo de si la planta tiene crecimiento primario o secundario, epidermis y peridermis, respectivamente.
Epidermis
Durante el crecimiento primario de la planta la epidermis constituye el tejido de protección de tallos, hojas, raíces, flores, frutos y semillas. Se acepta que no existe en la caliptra de la raíz y que no está diferenciada en los meristemos apicales. Se origina a partir de la capa más externa del meristemo apical, también denominada protodermis. Se considera como tejido de protección ya que esa es una de sus funciones.
La epidermis está formada comúnmente por una sola fial de células, salvo algunas excepciones donde se aprecian disposiciones estratificadas, como es el caso de las raíces aéreas o de determinadas hojas como en las adelfas.

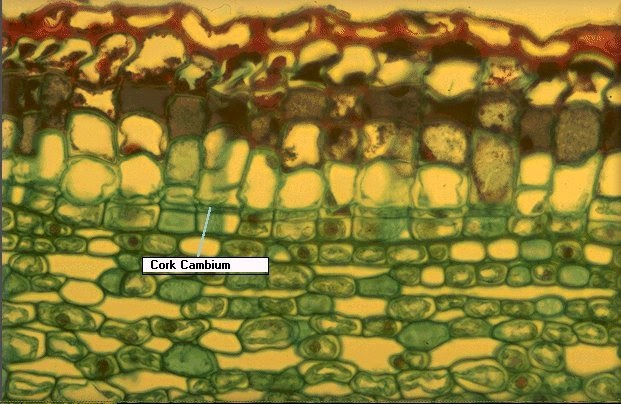
Peridermis
Peridermis
Se forma en aquellas partes de la raíz y del tallo que presentan crecimiento secundario, por la actividad del cambium suberoso o felógeno. Este meristemo secundario se origina de la desdiferenciación de las células parenquimáticas o colenquimáticas que se encuentran debajo de la epidermis, con lo que puede formar un cilindro completo o incompleto.
Tejidos glandulares
En las plantas es difícil discriminar entre excreción y secreción, puesto que los productos de ambos procesos suelen acumularse en los mismos compartimentos vegetales o en la superficie exterior de la planta. Se puede decir que la secreción, en términos generales, es la acumulación de metabolitos secundarios que no van a ser utilizados y de metabolitos primarios que van a ser utilizados de nuevo por la célula. Las células secretoras proceden de la diferenciación de otras células pertenecientes principalmente a la epidermis o al tejido parenquimático y no constituyen verdaderos tejidos.
Estructuras de secreción externas.
Existen multitud de estructuras secretoras en la superficie de la planta que se pueden encontrar formando pelos unicelulares o pluricelulares en la epidermis, o formando parte de la superficie epidérmica. Se originan por diferenciación y división de células epidérmicas.
Las estructuras secretoras en forma de pelo presentan generalmente una morfología constituida por un pié o pedúnculo y una cabeza que puede ser unicelular o pluricelular y que almacena el producto de secreción. Este producto es liberado mediante permeabilidad, a través de poros o por rotura de la cabeza del pelo.
Los hidatodos son estructuras que liberan agua, principalmente en las hojas, y que se acumula en forma de gotas. Este agua proviene directamente desde las traqueidas del xilema.
Los nectarios son estructuras secretoras productoras de azúcares resultantes de los elementos aportados por el floema. Se encuentran sobre todo en las flores, denominados nectarios florales, pero también en las partes vegetativas o nectarios extraflorales.
Los osmóforos son estructuras secretoras que producen el olor de las plantas mediante secreción de aceites volátiles. Las glándulas de la sal de las plantas halófitas secretan iones para regular el contenido en sales de la planta. Los tricomas de plantas urticantes secretan un líquido que produce irritación cutánea y que utilizan como mecanismo de defensa.
Estructuras de secreción interna
Las secreciones internas son productos que se almacenarán en el interior de los tejidos de la planta, a veces durante toda la vida de ésta. Las estructuras secretoras internas se encuentran alejadas de la epidermis y principalmente distribuidas en el parénquima cortical. Podemos distinguir tres grupos:
Las células secretoras. Son células aisladas que se diferencian de las células vecinas por su morfología, que puede variar desde formas isodiamétricas hasta tubos más o menos alargados. Sintetizan una amplia variedad de productos que almacenan en su interior, como resinas, mucílagos, taninos e incluso sustancias cristalizables.
Las cavidades y los conductos secretores se diferencian de los idioblastos en que su producto de secreción se acumula en los espacios intercelulares.
Por una separación de las células que resulta en un espacio central revestido por células secretoras, denominados espacios esquizogénicos, como es el caso de los conductos resiníferos. Por degradación de células que previamente habían sintetizado productos y éstos quedan en el hueco que dejan las células muertas, denominados espacios lisogénicos, este es el caso de las cavidades lisogénicas de los cítricos.
Conductos resiníferos

Los laticíferos son células individuales o en grupo que acumulan un líquido llamado látex. Cuando están formados por varias células se pueden organizar formando tubos.

Canales resiníferos del tallo. 


{kind=link}